University of Ličge,
Pathology Tower (BC-3), Sart-Tilman, B-4000 Ličge, BELGIUM
Phone : +32-4-366.72.75
Fax : +32-4-366.29.36
|
|
University of Ličge, Pathology Tower (BC-3), Sart-Tilman, B-4000 Ličge, BELGIUM Phone : +32-4-366.72.75 Fax : +32-4-366.29.36 |
Résumé de la thčse de doctorat en Sciences Biomédicales de Vincent LAMBERT défendue le 8 juillet 2005 ŕ l'Université de Ličge, Belgique
Ce travail a été réalisé au sein du
Dans le premier chapitre du travail consacré ŕ l’étude de l’intervention des protéases ŕ sérine dans la néovascularisation choroďdienne, nous avons montré que :
- PAI-1, uPA, tPA et uPAR sont exprimés ŕ la fois dans les membranes néovasculaires humaines et expérimentales;
- une protéolyse excessive (par absence de PAI-1) ou déficiente (par excčs de PAI-1, ou par inactivation de uPA, tPA ou Plg) empęchent toutes deux le développement de la réaction néovasculaire dans la choroďde;
- l’effet de PAI-1 sur le processus néovasculaire est pro-angiogčne ou anti-angiogčne en fonction de la dose étudiée, et s’exerce par le contrôle de l’activité fibrinolytique d’avantage qu’au travers des interactions avec la vitronectine;
- l’absence d’uPA, tPA ou Plg entraîne une accumulation de fibrinogčne/fibrine dans les lésions induites par le laser.
Le rôle essentiel joué par PAI-1 dans l’angiogenčse pathologique choroďdienne peut s’expliquer par le caractčre multifonctionnel de cette molécule. Bien que les mécanismes d’action de PAI-1 restent ŕ préciser, au moins deux théories peuvent ętre proposées. La premičre suggčre qu’en protégeant la matrice extra-cellulaire d’une dégradation excessive, PAI-1 permettrait de stabiliser la matrice, qui agit tel un échafaudage nécessaire ŕ la migration des cellules endothéliales et ŕ l’assemblage de ces cellules endothéliales en capillaires [Pepper and Montesano, 1990]. Le retard d’angiogenčse, observé en absence de plasminogčne, conforte également l’importance d’un contrôle précis de la protéolyse médiée par la plasmine [Bajou et al., 2001]. De plus, la séquestration de PAI-1 au sein de la matrice extra-cellulaire est compatible avec un tel rôle. Donc, un équilibre précis entre les enzymes protéolytiques et leurs inhibiteurs semble ętre essentiel ŕ la migration des cellules endothéliales et ŕ leur différenciation en vaisseaux fonctionnels. Dans un modčle d’anneaux d’aortes, aucun vaisseau n’a été formé ŕ partir d’animaux déficients pour le PAI-1. L’ajout de PAI-1 recombinant exogčne, ŕ des concentrations physiologiques, permet de nouveaux le bourgeonnement des micro-vaisseaux, tandis que l’usage de concentrations dites thérapeutiques inhibe l’angiogenčse [Devy et al., 2002]. De façon similaire, le PAI-1 inhibe l’angiogenčse induite par le bFGF, lorsqu’il est employé ŕ fortes concentrations [Stefansson et al., 2001]. PAI-1 inhibe également l’angiogenčse pathologique dans un modčle de rétinopathie du prématuré chez le rat [Penn and Rajaratnam, 2003]. Le PAI-1 semble donc avoir une dualité de fonction arborant une activité pro-angiogénique ŕ des concentrations physiologiques, et des effets anti-angiogéniques ŕ de hautes concentrations dites pharmacologiques[Bajou et al., 2004b].
Le second mécanisme d’action de PAI-1 durant l’angiogenčse peut ętre mis en relation avec sa capacité ŕ interagir avec l’uPAR, les intégrines, et la vitronectine, et, par conséquent, de contrôler la migration cellulaire. Lorsque le PAI-1 est lié ŕ la vitronectine [Zhou et al., 2003;Czekay et al., 2003], le site d’adhésion des intégrines devient cryptique et l’adhésion cellulaire est bloquée. Toutefois, l’expression de l’uPA compense l’effet d’anti-adhésion du PAI-1, en déstabilisant l’interaction entre la vitronectine et le PAI-1. La liaison de l’uPAR et de la vitronectine pourra, dčs lors, avoir cours ŕ nouveau et permettra l’accčs des intégrines cellulaires sur les sites d’adhésion RGD. En tenant compte du concept selon lequel la migration cellulaire est considérée comme une succession d’attachements et de détachements, l’effet du PAI-1 pourrait donc ętre vu comme de type migratoire [Stefansson and Lawrence, 2003;Czekay and Loskutoff, 2004;Degryse et al., 2004].
Afin d’élucider le mécanisme d’action de PAI-1, des adénovirus ont été utilisés dans le but de délivrer deux formes mutantes de PAI-1 ŕ des souris déficientes pour le PAI-1, dans le modčle de transplantation cité plus haut [Bajou et al., 2001] et, dans un modčle d’anneaux aortiques [Devy et al., 2002]. Les PAI-1 mutants, soit ont gardé une liaison normale ŕ la vitronectine, mais ne peuvent plus inhiber l’uPA ou le tPA, soit inhibent normalement les activateurs du plasminogčne, mais ont une liaison sur la vitronectine négligeable. Dans les deux modčles, aussi bien in vivo, qu’in vitro, l’angiogenčse est restaurée uniquement par le PAI-1 mutant ayant conservé son activité inhibitrice des activateurs du plasminogčne intacte, démontrant que le besoin de PAI-1 est lié ŕ sa capacité d’inhiber une protéolyse excessive plutôt que d’interagir avec la vitronectine. En accord avec ces données, l’angiogenčse et l’invasion tumorale ont été aussi bien observées chez des souris déficientes pour la vitronectine que dans le groupe témoin [Bajou et al., 2001]. Dans une étude séparée utilisant les PAI-1 mutants, l’inhibition de l’angiogenčse induite par le bFGF dans un modčle de membrane chorďo-allantoďde d’embryon de poulet requiert aussi bien l’activité anti-protéinasique que la capacité de liaison ŕ la vitronectine du PAI-1 [Stefansson et al., 2001]. Toutes ces données suggčrent que, suivant la situation, PAI-1 peut contrôler l’angiogenčse en régulant les évčnements protéolytiques et/ou non-protéolytiques dans la migration des cellules endothéliales [Stefansson et al., 2003].
Le corollaire de ces recherches est que l’utilisation de PAI-1 en tant que cible thérapeutique peut s’avérer hasardeuse et dangereuse. Nos résultats peuvent en partie expliquer l’efficacité dose-dépendante de médicaments, tel que l’anécortave acétate, et mettre en garde contre des stratégies pharmacologiques incontrôlées dans lesquelles les agonistes et antagonistes de PAI-1 sont utilisés pour l’inhibition de la néovascularisation choroďdienne et pour lesquelles existe une courbe biphasique dose-réponse.
Dans le chapitre consacré ŕ l’étude de l’influence des métalloprotéinases, nous montrons que :
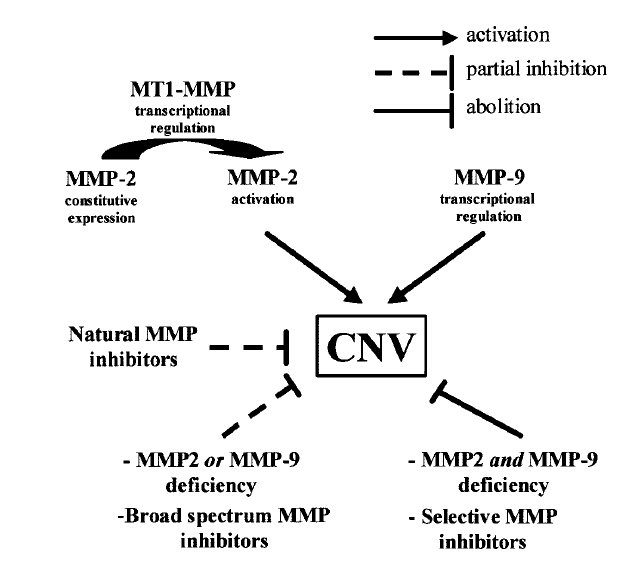
- le profil d’apparition et d’expression de la MMP-9 dans la lésion ressemble au profil observé au cours de la cicatrisation et coďncide avec l'arrivée des monocytes dans la lésion;
- l’absence de MMP-9 réduit l’importance de la réaction néovasculaire;
- l’inactivation combinée de la MMP-2 et de la MMP-9 prévient le développement de la néovascularisation choroďdienne plus efficacement que la déficience isolée d’un des gčnes;
- l'activation du zymogčne pro-MMP-2 est la conséquence d'une surexpression de MT1-MMP;
- un inhibiteur naturel ou synthétique de métalloprotéinases peut prévenir ou entraîner une régression partielle des néovaisseaux choroďdiens.
Nous avons ainsi montré qu’une modulation contrôlée de l’activité protéolytique pouvait mener ŕ l’inhibition de l’angiogenčse. Un traitement expérimental utilisant un dérivé innovant, inhibiteur sélectif de l’activité de certaines MMPs [Maquoi et al., 2004], s’est montré particuličrement efficace dans notre modčle. Ces observations ouvrent donc la voie ŕ une stratégie combinant des molécules intervenant ŕ différentes étapes de la néovascularisation choroďdienne.
En conclusion, nos travaux pourront ouvrir de nouvelles perspectives thérapeutiques, en association avec d’autres traitements et de nouvelles technologies, dans le cadre de la lutte contre la dégénérescence maculaire liée ŕ l’âge aussi bien sur le plan curatif que préventif.